
Análise de campos microscópicos de culturas de HeLa e MDA-MB-231 após irradiação
em cinética temporal
Celso Vieira de Lima e Tarcisio Passos Ribeiro de Campos
Programa de Pós-graduação em Ciências e Técnicas Nucleares Departamento de Engenharia Nuclear,
Universidade Federal de Minas Gerais - Brasil
Dezembro de 2019
Resumo
O presente artigo estuda as alterações radioinduzidas em células neoplásicas de mama, da linhagem MDA-MB231, e de útero HeLa, que foram submetidas a raios X de 4MeV com dose única de 5Gy, em cinética de tempo de 2, 6, 12, 24-48 e 48-72 horas após irradiação. As células foram cultivadas em meio de cultura convencional, 10% de soro fetal bovino (SBF), em condições estéreis, mantidas em estufa climatizada com substituição do meio a cada dois dias até 80% de confluência, com concentração celular de 10ˆ6cel./cmˆ2 nos frascos de cultura T-25 e T-50, condições consideradas ideais para este ensaio. Dois frascos, idênticos aos anteriores, foram mantidos livres da radiação para controle.
Os estudos microscópicos, mostram alterações na morfologia em diversas situações: em divisão celular, aderidas e desatachadas, fragmentos celulares (debris), pacotes apoptóticos, com vacúolos, onde os conjuntos celulares apresentam-se viáveis, ou em processo de apoptose e necrose.
Mostram ainda efeitos intensos na morfologia celular das células HeLa, após a irradiação, não resistindo estas por mais de 48 horas. Já as células MDA-MB-231 também sofreram mudanças morfológicas em menor escala, se adaptando ao tratamento e voltando ao ciclo celular em 72h com atividades superiores ao controle. Esses achados descrevem os efeitos da radiosensibilidade da linhagem HeLa e da radiorresistência da MDA-MB-231.
Palavras-chave: Morfologia celular, irradiação, células neoplásicas, imagem em microscopia invertida.
Introdução
A alta complexidade do corpo humano, em grande parte, está relacionada à diversidade celular e à especificidade tecidual no que se refere aos recursos de replicação celular (ANDRADE, et al., 2005). No contexto genômico, a multiplicação celular se dá de duas maneiras: Uma reducional, quando as células iniciais dão origem a células filhas com metade de seus cromossomos, encontrada em células gaméticas para produção dos gametas. Outra conservativa, processo encontrado em células somáticas que geram células filhas com a mesma quantidade de cromossomos. (KNISELY, et al., 2009).
Não se pode negar que os dois processos de divisão celular têm manifestações que as alinham em termos gerais, como a fase inicial da divisão celular onde todo o conteúdo celular é duplicado, período denominado de interfase, e a fase final da divisão com a diferenciação celular, quando são originados os gametas ou as células específicas de cada tecido (KNISELY, 2009; LODISH, 2000; BECKER, 2009; ALBERTS, 2002).
Uma vez ocorrida à diferenciação celular, as células vão multiplicar-se por um mecanismo chamado de hiperplasia induzido por estímulos tróficos, em especial pelo hormônio do crescimento (GH) liberado pela glândula adeno-hipófise (RABAJANTE, J.F.; BARBIERRA, A.L, 2015). Tão logo o indivíduo chegue à puberdade, com o início da vida reprodutiva, os níveis plasmáticos do GH são reduzidos e causam uma modificação no fenômeno de crescimento corporal. Neste momento, a hipertrofia representada pelo aumento do volume celular, que até então desempenhava uma função secundária, assume na puberdade uma posição de destaque sobre a hiperplasia, perdurando até o início do envelhecimento onde a atrofia celular vem substituir a hipertrofia ( GUILAK, F., 2000; GURUNDUTA, G.U., 2012).
Existem atualmente várias explicações na literatura para o processo da diferenciação celular, quais sejam: (i) ação hormonal do hormônio do crescimento (GH), do hormônio folículo estimulante (FSH) e do hormônio luteinizante (LH), (ii) controle da transcrição, (iii) processamento de RNA, (iv) controle do transporte do RNA, e (v) controle traducional (GURUNDUTA, G.U. e outros, 2012), no entanto, o entendimento sobre este evento ainda não está consolidado. Existem ainda vários fenômenos não explicados, como o efeito do microambiente celular sobre esta diferenciação, e mesmo os fatores que estão relacionados a falhas neste processo de diferenciação que tem como reflexo o surgimento das neoplasias (GURUNDUTA, G.U., 2012 ; HARRIS, J., 1997).
No entanto os estudos de diferenciação celular são feitos ao longo de uma métrica temporal, para que as ações celulares possam ser consolidadas e os aspectos da dinâmica das alterações morfológicas em campos microscópicos devidamente efetivados. Assim, existem vários aspectos ainda obscuros em relação à cinética temporal da diferenciação celular, entre eles o efeito das radiações ionizantes sobre esta, ou a variação de expressão das proteínas em virtude das alterações e/ou mutações genéticas (HADDAD, N., 2001; SILVA,M.B., 2001).
Vê-se que a literatura carece de estudos comparativos morfológicos em cinética temporal, sobre as alterações da diferenciação celular em células em divisão celular nos diversos tecidos humanos. O modelo clonogênico neoplásico pode ser empregado para tais estudos.
O presente artigo tem como objetivo qualificar as diversas apresentações morfológicas de células cancerosas em campos microscópicos, expostas a agentes exógenos como radiação, comparativas ao controle, em cinética temporal (SCHWARTSMANN, G.2001; JONES, H.W.2001; VASCONCELOS, A.B. 2001; WAY, L.W.2001).
Materiais e métodos
Linhagens cultivadas
Neste estudo foram utilizadas linhagens de células HeLa e MDAMB-231 estabelecidas em cultura, doadas pela Fundação Ezequiel Dias - FUNED e Instituto de Ciencias Biológicas da Universidade Federal de Minas Gerais - ICB/UFMG.
As células foram cultivadas em frascos T-25 e T-50 em meio convencional, RPMI-1640 (Sigma), acrescido de 10% de soro fetal bovino (SFB) como indutor de crescimento, e em condições estéreis.
A cultura foi mantida em estufa climatizada contendo CO2 a 37oC, com substituição do meio a cada 2 dias, até atingirem 100% de confluência, observada por microscopia óptica, com uma concentração da ordem de 106cél.cm-2 no frasco T-25 e T-50, considerando essas como as condições ideais para o ensaio.
Para realização do trabalho, dois grupos de material biológico foram estabelecidos. Um grupo controle e um grupo de estudo, ambos em duplicata para cada uma das duas linhagens estabelecidas. O grupo de estudo foi submetido a uma dose única de 5Gy, em acelerador linear de 4MV, por aproximadamente 4min. Em seguida os frascos contendo as células irradiadas retornaram para a estufa nas condições anteriores, por 2h para estabilização.
Resultados
Grupo controle
Nos frascos de controle estão células neoplásicas, não irradiadas, cuja configuração pode ser vista nas imagens da Fig.1. Na Fig.1A são identificadas células HeLa em condições ideais de cultivo em contato com células vizinhas (seta 1), e na Fig.1B, células MDAMB231 de forma irregular (estrelada) em condições ideais de cultivo (seta 4). Poucas células, em processo de descolamento do fundo do frasco, são observadas e são identificadas pela forma esférica e contraste acentuado (setas 2).
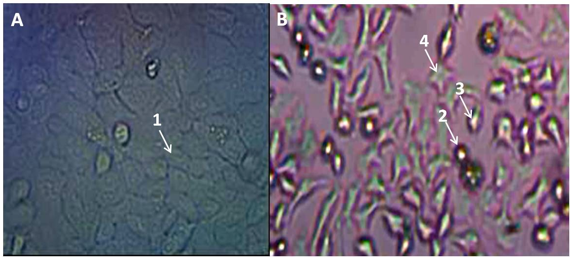
Fig. 1 - Fotografias de campos microscópicos de linhagens confluentes de células (A) HELA e (B) MDAMB,
controles, onde setas apontam distintas morfologias celulares nos campos.
Grupo de estudo
Nos frascos de estudo estão células neoplásicas irradiadas como descrito anteriormente. Até duas horas após a exposição, as imagens microscópicas não demonstram alterações significaivas no quesito morfológico das culturas de ambas as linhagens celulares.
Primeira observação: 6h pós-irradiação
Após a sexta hora pode-se ver, na Fig.2, ligeira displasia nas células HeLa com consequente descolamento do frasco.
Em relação aos Campos Microscópicos Celulares (CMC) da linhagem MDA-MB-231 observa-se presença de debris de células e células descoladas da garrafa, células em apoptose, visto que possuíam grandes vacúolos (identificados como micro esferas translúcidas intracelulares), o fenômeno da autólise, característico da apoptose. Observa-se também campos contendo células com modificações estruturais (displasias), e células com morfologia normal. Os CMC apresentam menor confluência que o controle.
As figuras 2A e 2B mostram células das linhagens HeLa e MDAMB-231 6h pós-irradiação. Pode-se identificar na Fig.2A, nos campos da célula HeLa unidades que estão em processo de descolamento do frasco em possível apoptose (seta 1), célula com morfologia diferenciada do padrão controle (seta 2) e Fig.2B, na linhagem MDA-MB-231 pode-se observar pacotes apoptóticos descolados que assumem geometria esférica (seta 3), e grandes vacúolos intracelulares (seta 4).

Fig.2 - Campos celulares de cultura de células (A) HeLa (B) MDAMB, 6 h pós irradiação de 5Gy.
Segunda observação: 12h pós-irradiação
Dose horas pós-irradiação, nos frascos de cultura de HeLa, observa-se através de CMC que cerca de 80% das células haviam sofrido apoptose e desprendidas do frasco, deixando poucos debris aderidos. Entretanto, observa-se também, células HeLa que permaneceram viáveis, possivelmente com ciclo celular interrompido, visto que não foram identificados crescimento do número de células bem como divisão celular.
Nos frascos de MDAMB-231 também foi observado lise; no entanto, em proporção menor, estimada em cerca de 40%. Foram observados também apoptoses com debris aderidos ao frasco. Os restantes das células permaneceram com morfologia diferente das células controle; entretanto possivelmente viáveis. Pode-se observar também ciclo celular interrompido neste período de tempo, considerando a ausência de células em divisão celular.
Cabe aqui a observação que as células HeLa após morte se desprendiam completamente do frasco de cultura, observado muito poucos debris aderidos, ao contrário do que foi identificado nos frascos com células MDA-MB-231, onde seus debris permaneciam aderidos.
Enquanto, 80% das células HeLa haviam sido lisadas ou sofrido apoptose tendo os fragmentos desprendidos, apenas 30% das células MDA-MB-231 sofreram o mesmo processo. Assim ficou claro que as células MDA-MB-231 são mais resistentes a radiação que as células HeLa nas mesmas condições de cultivo e irradiação. As figuras 3A, 3B e 3C mostram fotografias das linhagens HeLa e MDA-MB-231 a 12h pós-irradiação. O CMC em HeLa é muito esparso onde podem ser fotografadas algumas células em displasia. Pode-se identificar na Fig.3A, ampliada em X800 fragmentos apoptóticos de células HeLa (seta 1 e 2), aglomerado de células em displasia (seta 2); Fig.3B (seta 3); e, Fig.3C células binucleadas (seta 4), células em desprendimento do frasco com geometria esférica (seta 5), célula com grande vacúolo (seta 6), fragmentos nucleares originados de apoptose (seta 7).

Fig. 3 - Campos microscópicos de culturas de celulares de (A, B) HeLa; e, (C) MDAMB-231após 12 h pós 5 Gy.
Terceira observação: 24-48h pós-irradiação.
Na Fig. 4A pode se identificar restos celulares (debris) da linhagem HeLA, que tomam a forma de estruturas esféricas deslocadas da superfície, nas indicações setas 1, 2 e 3. Pode-se identificar na Fig.4A (seta 3) uma célula em necrose.
Por sua vez, na Fig.4B, identifica-se uma pluralidade de efeitos e ações celulares. Assim, na Fig4B (seta 4) são identificados células viáveis (estreladas) em seu ciclo celular sem possíveis alterações. Observam-se também células viáveis; no entanto, apresentando um alongamento quase na forma filamentar do seu citoplasma (seta 5). É possível identificar restos celulares oriundos do tratamento com a radiação (seta 6). E em geral em todo campo, exemplificado Fig.4B (seta 7), há um grande número e células esféricas descoladas ainda indefinidas em relação ao retorno ao ciclo celular ou a entrada na apoptose ou necrose.

Fig.4 - Campos microscópicos de culturas de celulares de (A) HeLa (B) MDAMB-231a 24-48h a 5Gy.
Quarta observação: 48-72h pós-irradiação
A análise dos campos microscópicos das linhagens de HeLa a 48-72h, mostra que as células HeLa não resistiram a radiação aguda de 5Gy, sofrendo apoptosis, necrosis e total desprendimentos dos fragmentos celulares para o meio, encerrando a análise. Enquanto, no frasco de células MDA-MB-231, houve ainda a identificação de células viáveis em transformações morfológicas. As figuras 5A e 5B mostram fotografias das linhagens MDAMB-231 pós 72h pós-irradiação, onde se destacam células viáveis morfologicamente diferentes do controle (seta 1), células em apoptoses em processo de desprendimento (seta 2), células esféricas aferidas no frasco (seta 3), células em processo intermediário entre morfologias indicadas em 1 e 2 (seta 4); enquanto na Fig.5B, identificam-se células viáveis com morfologia diferenciada em relação ao controle (estrelada) onde há a presença de filamentos alongados (setas 5 e 6). Já na Fig.5C é mostrado CMC com conjunto de células em crescimento acelerado com produção de uma segunda camada sobrepondo o tapete celular (setas 7, 8 e 9).
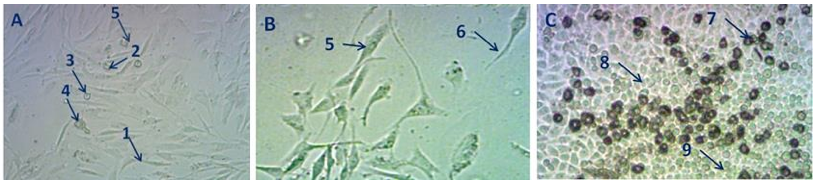
Fig.5 - Campos microscópicos de culturas de celulares de MDAMB-231,48- 72 h pós-irradiação com doseabsorvida de 5 Gy,
indicativo de crescimento sobre os debris de células lisadas.
DISCUSSÃO
Após irradiação, existem várias condições que caracterizam os fenótipos celulares visibilizados em CMC no frasco experimental: células viáveis irradiadas; viáveis não irradiadas; debris resultante da lise celular de células irradiadas não viáveis; células em processo apoptótico; ou em mitose; e, viáveis fora do clico celular, coexistindo no mesmo frasco. A existência de células viáveis não irradiadas consiste da probabilidade de ocorrência, em condições estocásticas, de células que não sofreram dano por radicais livres produzidos no processo de exposição a 5Gy, coexistem em conjunto com os outros tipos de fenótipos. Desta forma, CMC apresentam uma diversidade muito grande de tipos de estágio celular. (ANDRADE, L. M., et al., 2005)
Após a décima segunda hora conforme já mencionado, observamos uma pequena população de células HeLa, que permaneceram aderidas até 48h em CMC esparsos. Um numero significativo de células HeLa sofreram o processo de lise. No entanto após 12h não foi observado um aumento no numero de HeLa o que pode indicar uma total inibição do ciclo celular.
As células HeLa sofreram apoptose e necrose no espaço temporal de 48-72h, mostrando ser a dose de 5Gy aguda efetiva para o total controle clonogênico desta linhagem. Não foi feita análise após 72h devido à ausência de células nos campos microscópicos.
Após 72h da exposição, notou-se uma recuperação numérica das células MDA-MB-231 que apresentou crescimento acelerado. Observou-se uma estagnação de crescimento das células HeLa, apresentando displasia e campos totalmente esparsos de células, novamente confirmando a observação que células HeLa são fortemente afetadas pela dose absorvida arbitrariamente aplicada, enquanto células MDA-MB-231 puderam se recuperar em condições favoráveis, possivelmente superior ao controle. Este achado pode ser confirmado em estudos anteriores onde houve a caracterização de um crescimento substancial das células MDA-MB-231 após 72h da exposição. (ANDRADE, L. M., et al., 2005). Este mesmo desenvolvimento foi observado em CMC após 72h para as células MDA-MB-231.
As células MDA-MB-231, no período de 72h, não apresentaram nenhum descolamento ou alteração de sua forma. No entanto após a 12ªh observamos que cerca de 30 a 40% das células MDA-MB-231 haviam sofrido apoptose ou desprendidas do frasco, permanecendo o restante aderido; entretanto em displasia diferente da morfologia de forma estrelada no controle. Dados anteriores de nosso laboratório apontam para comportamento semelhante, corroborando nossos dados para estabelecer nestas linhagens um padrão funcional.
Semelhante à linhagem HeLa observadas a 6h, as células MDA-MB-231, permaneceram aderidas ao frasco de cultura a 12h com forma displasica, sem presença de células em mitose que sugere inibição radioinduzida do ciclo celular. No entanto, ao contrário das células HeLa, após a 48ªh observou-se nas MDA-MB-231 início de divisão celular, que foi intensificando nas próximas 24h tendo seu ápice por volta de 72h após a irradiação. Tal crescimento foi considerado superior a evolução natural de confluência da linhagem em condições ótimas de cultivo.
Estes achados também foram observados na literatura, com irradiação das mesmas células na dose de 50Gy usando o ensaio de viabilidade celular por MTT. (ANDRADE, L. M., et al., 2005) Foram observados a inibição do ciclo celular até o período de 48h e o retorno do crescimento após as 72h de irradiação, sendo este crescimento mensurado em MTT 50% maior que o crescimento controle. (ANDRADE, L. M., et al., 2005) Este fato também foi observado nestes ensaios.
A opção de dose absorvida foi de 5Gy em fração única, 10 vezes menor que a total dose terapêutica de 50Gy aplicada de forma fracionada de 2Gy em tratamento convencional de câncer de mama, através de portais opostos paralelos. Em nossos ensaios foram observados a não letalidade da dose nas duas primeiras horas, fato que não se manteve até a sexta hora onde houve perda celular estimada nos CMC de 30 a 40% das células MDA-MB-231 e cerca de 80% das células HeLa, mostrando ser esta última uma linhagem celular mais sensível à radiação nas condições experimentais, visto ter sido eliminada completamente até 72h após a irradiação com a dose preconizada. Já as células MDA-MB-231, após a 12ª hora, estabilizou-se permanecendo inerte até a 48ªh onde reiniciou o processo de clonogenicidade de forma estimada superior ao controle.
No entanto apesar de uma sensibilidade diferenciada para as duas linhagens celulares um comportamento comum chama a atenção, o fato do ciclo celular (divisão celular), permanecer inibido até 48 horas após a irradiação em ambas as linhagens, acompanhado por perdas celulares por apoptosis e necrosis. Após este período, a linhagem MDA-MB-231 experimenta um crescimento celular em taxas superiores ao controle. Neste sentido, as observações sugerem a existência de um mecanismo de contra-regulação contrapondo ao processo na fase inicial que leva ao bloqueio do ciclo celular observado até 48h. Este mecanismo está relacionado ao tempo necessário para uma possível reparação das células irradiadas, seguida pelo disparo do crescimento com seu ápice em 72h após a irradiação. Os dados ainda sugerem a possibilidade da ação após a irradiação de “fatores inibitórios” e “fatores excitatórios” radioinduzidos do crescimento celular que estariam atuando em um mecanismo de contra regulação modulando o ciclo celular, fato em nenhum momento observado no ensaio controle. Ainda pela amplitude das respostas, podemos observar um potente efeito inibitório e excitatório nas células irradiadas MDA-MB-231, em cultura. Fenômeno que pode também ser observado in vivo dependendo da linhagem neoplásica. Novos estudos estão sendo feitos pelos autores no sentido de esclarecer este mecanismo, identificando as substâncias indutoras e inibidoras do crescimento, na busca do entendimento do processo de instalação e evolução de clones cancerosos, com possibilidades de se tornar um instrumento de prognóstico e terapia.
Conclusão
Os resultados encontrados, sugerem que células da linhagem neoplásica de útero HeLa, são mais sensíveis ao tratamento com a radiação utilizada, pois não demonstrou capacidade de reparar as lesões celulares causadas pela radiação, sendo a cultura praticamente extinta 48h após exposição de 5Gy. Ao contrário, as células do câncer de mama da linhagem MDA-MB-231, apresentaram-se resistentes à dose de 5Gy, com aparente sensibilidade nas primeiras 12h, mas com possível retorno a clonogenicidade após 72h. com excessivo processo clonogênico, superior ao encontrado no controle. Assim como o observado, existem linhagens mais radio-resistentes que outras, e que o tratamento utilizado de maneira convencional deve levar em conta estas resistências celulares específicas na montagem dos protocolos de radioterapia.
Agradecimentos
Os autores agradecem ao Conselho Nacional de Desenvolvimento da Ciência e Tecnologia (CNPq), ao apoio financeiro do processo 456719/2013-0, ao Conselho de Apoio à pós-Graduação e Ensino Superior (CAPES) pela bolsa de estudos PNPD concedida.
Referências bibliográficas
ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K.; WALTER, P. Molecular Biology of the Cell. 4th edition. New York: Garland Science. 2002.
ANDRADE, L.M.; LEITE, M.F.; GOES, A.M.; CAMPOS, T.P.R. Cellular Viability and Apoptosis of Human Breast Cancer MDAMB-231 Cell Line After Co60 Irradiation. Braz. arch. biol. technol. 2005.
BECKER, W.M.; KLEINSMITH, L.J.; HARDIN, J.; & BERTONI, G.P. The World of the Cell (17th edition). Pearson Education, Inc. 2009.
FREDRIKSSON, I.; LILJERGREN, G.; ARNESON, L.G. time trends in the results of breast conservation in 4694 women. Eur.J.Cancer. 2001.
GUILAK, F.; COHEN, D. M..; ESTES, B. T.; GIMBLE, J. M.; LIEDTKE, W.; CHEN, C.S. "Control of Stem Cell Fate by Physical Interactions with the Extracellular Matrix"
GURUNDUTA, G.U.; NEERAJ, K.S.; VIMAL, K.S.; YOGESH, K.V.; PALLAVI, G.; TRIPATHI, R.P. Stem cell therapy: A novel & futuristic treatment modality for disaster injuries. Indian J Med Res. 2012.
HADDAD, N.; SILVA, M.B. Mortalidade por Neoplasias em Mulheres em Idade Reprodutiva-15 a 49 anos- no Estado de São Paulo, Brasil, de 1991 a 1995. Rev.Ass.Med.Brasil. 2001.
HARRIS, J.; MORROW, M.; NORTON, L. Malignant Tumors of the Breast. In Cancer, Principles & Practice of Oncology. Lippincott-Raven, Philadelphia. 1997.
JONES,H. W.; WENTS, A.C.; BURNETT, L.S.N. Tratado de Ginecologia, Guanabara Koogan, Rio de Janeiro. 2001.
KNISELY, K.; GILBERT, S.F. Developmental Biology (8th ed.). Sunderland, Mass: Sinauer Associates. 2009.
LODISH, H. Mollecular Cell Biology (4th ed.).New York: W.H. Freeman. 2000.
RABAJANTE, J.F.; BABIERRA, A.L. Branching and oscillations in the epigenetic landscape of cell-fate determination. 2015.
ROBBINS, S.L.; COTRAN, R.S.; KUMAR, V. R. Pathologic Basis of Disease, W.B Saunders Company, Philadelphia. 1989.
SCHWARTSMANN, G. Breast cancer in South America: challenges to improve early detection and medical management of a public health problem. J. Clin. Oncol. 2001.
TESSARO,S.; BÉRIA, J.U.; TOMASI, E. Contraceptivos Orais e Câncer de Mama: estudo de casos e controles. Rev. Saúde Pub. 2001
VASCONCELOS, A.B.; MENDONÇA, G.A.; SICHIERI, R. H. weight, weight change and risk of breast cancer in Rio de Janeiro, Brasil. São Paulo Med. J. 2001.
WAY, L.W. Current surgical Diagnosis & Treatment, Appleton Lange.1991.